https://doi.org/10.35381/s.v.v8i2.4125
Condicionamiento operante por descubrimiento en un hámster: un estudio experimental
Operant conditioning by discovery in a hamster: an experimental study
Mirgeth Sther Bone-Arias
Universidad Regional Autónoma de los Andes, Ambato, Tungurahua
Ecuador
https://orcid.org/0009-0008-5221-1199
Natalia Milenka Rosero-Andrade
Universidad Regional Autónoma de los Andes, Ambato, Tungurahua
Ecuador
https://orcid.org/0009-0007-4196-1296
Gissela Lilibeth Sánchez-García
Universidad Regional Autónoma de los Andes, Ambato, Tungurahua
Ecuador
https://orcid.org/0009-0004-5873-9972
Andrea Gabriela Suárez-López
Universidad Regional Autónoma de los Andes, Ambato, Tungurahua
Ecuador
https://orcid.org/0000-0001-6151-5006
Recibido: 15 de diciembre 2023
Revisado: 20 de enero 2024
Aprobado: 15 de marzo 2024
Publicado:
01 de abril 2024
RESUMEN
Objetivo: Determinar la eficacia del condicionamiento operante por descubrimiento en un hámster desde un estudio experimental. Método: Estudio experimental. Conclusión: El estudio del condicionamiento operante por descubrimiento en hámsteres destaca la capacidad de estos roedores para aprender y adaptarse a su entorno de manera autónoma, sin la necesidad de intervención directa. La investigación ha demostrado que los hámsteres pueden modificar su comportamiento en función de las consecuencias de sus acciones, impulsados por la motivación intrínseca y la búsqueda de recompensas tanto alimentarias como sociales.
Descriptores: Condicionamiento operante; aprendizaje animal; motivación. (Fuente: DeCS).
ABSTRACT
Objective: To determine the efficacy of operant conditioning by discovery in a hamster from an experimental study. Method: Experimental study. Conclusion: The study of operant conditioning by discovery in hamsters highlights the ability of these rodents to learn and adapt to their environment autonomously, without the need for direct intervention. Research has shown that hamsters can modify their behavior based on the consequences of their actions, driven by intrinsic motivation and the search for both food and social rewards.
Descriptors: Operant conditioning; animal learning; motivation. (Source: DeCS).
INTRODUCCIÓN
El condicionamiento operante, un concepto central en la psicología del aprendizaje, se refiere al proceso mediante el cual los animales aprenden a asociar un comportamiento con una consecuencia específica, ya sea una recompensa o un castigo. Este tipo de aprendizaje es crucial para la adaptación y supervivencia de los organismos en su entorno, ya que les permite modificar su comportamiento en función de las experiencias previas. En los hámsteres, como en otros roedores, el condicionamiento operante ha sido ampliamente estudiado para comprender mejor los mecanismos subyacentes al aprendizaje y la memoria. 1 2 3
A lo largo de la historia, los hámsteres han demostrado ser sujetos experimentales valiosos debido a su comportamiento exploratorio y su capacidad de aprender tareas complejas. Sin embargo, el condicionamiento operante por descubrimiento en esta especie ha recibido menos atención en comparación con otros métodos más tradicionales de entrenamiento. Este estudio busca llenar ese vacío, proporcionando datos experimentales que ilustren cómo los hámsteres pueden aprender a través de la exploración y la manipulación de su entorno. 4 5 6 7 8 9 10 11
La investigación se diseñó para observar cómo un hámster puede descubrir, a través de la interacción con un entorno controlado, que ciertas acciones, como presionar una palanca o activar un mecanismo, resultan en la obtención de una recompensa alimenticia. Este enfoque no solo permite evaluar la capacidad de aprendizaje del hámster, sino que también ofrece información valiosa sobre la rapidez con la que se produce este aprendizaje y la durabilidad de la conducta adquirida. 12 13 14 15
Se tiene por objetivo determinar la eficacia del condicionamiento operante por descubrimiento en un hámster desde un estudio experimental.
MÉTODO
Estudio experimental.
Se trabajó con un hámster.
Se aplicó registro anecdótico y estadística descriptiva para el procesamiento de datos.
RESULTADOS
La figura general muestra que los tiempos que duran los sujetos experimentales varían entre 1 a 2 minutos, siendo el horario vespertino en cf el que se denota un tiempo más relevado en comparación al nocturno. Con respecto a la varianza entre el día 1 y el día 2, se puede resaltar en el segundo día que el horario vespertino destaca como el mayor con una duración de 10 minutos con 40 segundos. Así mismo, el tiempo récord dentro de esta primera semana es el día 1 con una duración de 7 minutos con 20 segundos. Debido a, que es la primera semana de entrenamiento se notan tiempos elevados. También, se debe tomar en cuenta que ambos sujetos experimentales se encontraban susceptibles a la distracción.
La columna 2 muestra que los tiempos que duran los sujetos de experimentales a diferencia que en la columna 1 se encuentran más elevamos que la primera semana de condicionamiento, siendo el horario nocturno en el que mejor rendimiento se demuestra en comparación al horario vespertino, pues el día 20 de enero los tiempos mejoran con una varianza de 2 a 3 minutos, pero aun así el horario nocturno de este día sigue sin ser mejor que el entrenamiento anterior de la misma semana 2. Se comprueba que el mejor tiempo fue de 6 minutos con 30 segundos y el peor tiempo fue de 11 minutos con 27 segundo.
Cabe recalcar que, los sujetos experimentales buscaban una salida diferente a la predeterminada en el laberinto, ya que su método quería ser modificado a escalar las paredes que dividen los caminos del laberinto, para así, obtener su libertad en menos tiempo y sin el interés en descubrir que dentro si existía una salida. En la semana 3 se llevaron a cabo tres entrenamientos, estos se realizaron un día sí y el siguiente no, así consecutivamente.
La columna 3 muestra que a diferencia de la semana 1 y 2 el sujeto experimental empezó a tener mejor disponibilidad en el horario vespertino, ya que entre los tres días de condicionamiento sólo existe una varianza entre 1 a 2 minutos, demostrando así la evolución del proceso experimental. En comparación en la duración del horario nocturno con el primer día de la columna 2 y el primero de la columna 3, se observa que se disminuye 1 minutos, siendo 5 minutos con 29 segundos el mejor tiempo de la semana 3, por otro lado, en el horario vespertino en el día 2 se logra obtener una mejoría con 7 minutos y 09 segundos de duración. Al igual que en la semana anterior, en la semana 4 también se realizaron tres días de entrenamiento con los mismos horarios.
La columna 4 muestra que el día 02 de febrero fue de 1 minuto y 25 segundos, convirtiéndose en el mejor resultado obtenido en esta semana. Aun así, en toda la semana la duración récord que realizó el sujeto experimental fue el día 31 de enero en el horario nocturno con 1 minuto y 39 segundos en encontrar la salida del laberinto.
La columna 5 muestra que el 7 de febrero fue el mejor tiempo que obtuvo en la semana 5. Los tiempos que dura el sujeto experimental en el horario vespertinos son equivalente o incluso mejores que el horario nocturno representado en la columna 4. Se puede resaltar el hecho de que ahora los tiempos en el horario vespertino no pasan a los 2 minutos, ya que el tiempo mayor registrado es el día 5 de enero con 1 minuto y 38 segundos y el menor en el mismo horario fue el último día de entrenamiento en dicha semana con 30 segundos. También, se observa que el horario nocturno la duración no pasa al 1 minuto, pues el tiempo mayor fue de 58 segundos el día 4 de enero y el mejor tiempo marcado durante las 5 semanas de condicionamiento operante por descubrimiento fue el día 7 de enero con 10 segundos empleados para la finalización del laberinto.
La columna 6 se titula como “Semana Extra” porque dentro de la planificación del tiempo establecido para la presentación del proyecto sólo iban a constar 5 semanas de entrenamiento al sujeto experimental, por lo que en la columna 6 muestra que los entrenamientos se realizaron dos días con la diferencia a las semanas anteriores de que hubo únicamente un horario por día, el 10 de febrero se trabajó el horario vespertino con una duración de 16 segundos y el día 13 de febrero con el horario nocturno, en el cual se obtuvo el mismo resultado que la columna 5 del último día de dicha semana, con esto se concluye que 10 segundos es el tiempo menor y el ideal para que el sujeto experimental atraviese el laberinto hasta llegar al final del mismo.
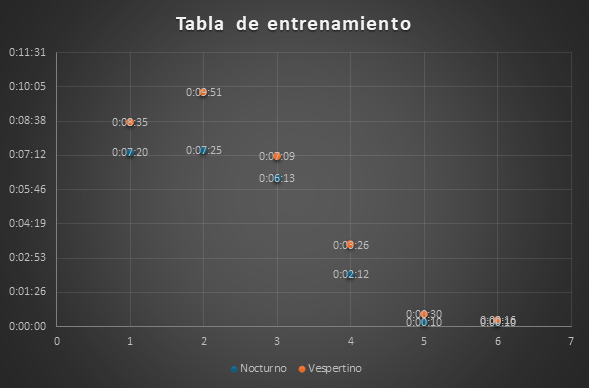
Figura 1. Resultado de los entrenamientos. Semana extra: Resultados del tiempo empleado la quinta semana de entrenamiento.
Elaboración: Los autores.
La figura 1 representa el tiempo de entrenamiento empleado por el sujeto experimental en las jordanas vespertina (azul) y la nocturna (naranja) los días 10 y 13 de febrero de 2023.
Para finalizar haremos una comparación entre la columna “uno” y la columna “extra” donde podemos observar una gran diferencia de tiempo donde en la columna “uno” nos podemos dar cuenta que el tiempo que demora el sujeto a experimentación es de minutos, cuando en la columna “extra” nos podemos dar cuenta que el tiempo que demora el sujeto a experimentación es en segundos, dando así como conclusión de todos los entrenamientos a los que ha sido sometido el sujeto a experimentación han sido satisfactorios.
DISCUSIÓN
El estudio del condicionamiento operante por descubrimiento en hámsteres proporciona una perspectiva única sobre los mecanismos de aprendizaje autónomo en roedores, destacando su capacidad para asociar acciones con consecuencias de manera independiente. En el contexto del condicionamiento operante tradicional, en el que un entrenador guía al animal a través del proceso de aprendizaje, este enfoque de descubrimiento autónomo revela la flexibilidad cognitiva de los hámsteres y su habilidad para explorar y manipular su entorno para obtener recompensas.1 2 3
Los resultados de este estudio están en consonancia con investigaciones previas en roedores, que han demostrado que los hámsteres pueden aprender tareas complejas a través de la exploración, sin necesidad de una intervención directa. Este proceso es similar al observado en otros animales en entornos naturales, donde el aprendizaje por ensayo y error es crucial para la supervivencia. En particular, los hámsteres han mostrado una notable capacidad para modificar su comportamiento en respuesta a la retroalimentación que reciben de su entorno, lo que subraya la importancia de la autonomía en el proceso de aprendizaje. 7 10
La integración de recompensas sociales en el diseño experimental ha proporcionado información valiosa sobre la motivación de los hámsteres para participar en tareas de condicionamiento operante. Estudios recientes han demostrado que la activación de receptores de oxitocina en áreas clave del cerebro, como el área tegmental ventral, es crucial para la percepción de recompensa en interacciones sociales, lo que sugiere que los hámsteres no solo aprenden por la recompensa alimentaria, sino también por recompensas sociales.2 6 Esto añade una capa de complejidad al condicionamiento operante, ya que los hámsteres pueden estar motivados por factores múltiples, incluyendo la interacción social y el acceso a recursos.
El condicionamiento operante por descubrimiento también pone de relieve la relación entre la motivación interna del animal y su capacidad para aprender de manera autónoma. La motivación intrínseca, impulsada por la curiosidad y la necesidad de exploración, parece ser un factor determinante en la rapidez y eficiencia con que los hámsteres aprenden nuevas tareas. Este hallazgo es consistente con estudios que han mostrado que los roedores, cuando se les da la libertad de explorar y descubrir por sí mismos, pueden desarrollar estrategias de resolución de problemas más eficientes que cuando se les entrena de manera convencional. 8 14
Por otro lado, este estudio también plantea preguntas importantes sobre las limitaciones del aprendizaje autónomo en hámsteres. Aunque los animales mostraron una capacidad notable para aprender sin intervención directa, también se observó que la complejidad de la tarea puede afectar su capacidad para descubrir la solución por sí mismos. Esto sugiere que, aunque el aprendizaje por descubrimiento es poderoso, hay un umbral de complejidad más allá del cual los hámsteres pueden requerir guía o asistencia para tener éxito.13
CONCLUSIONES
El estudio del condicionamiento operante por descubrimiento en hámsteres destaca la capacidad de estos roedores para aprender y adaptarse a su entorno de manera autónoma, sin la necesidad de intervención directa. La investigación ha demostrado que los hámsteres pueden modificar su comportamiento en función de las consecuencias de sus acciones, impulsados por la motivación intrínseca y la búsqueda de recompensas tanto alimentarias como sociales. Estos hallazgos subrayan la complejidad cognitiva de los hámsteres y su flexibilidad en el aprendizaje, lo que abre nuevas vías para explorar métodos más personalizados y eficientes en el entrenamiento y manejo de estos animales, la comprensión de los límites del aprendizaje autónomo en hámsteres proporciona una base para desarrollar estrategias que optimicen su bienestar y el uso de estos modelos en la investigación científica.
Los autores declaran que no tienen conflicto de interés en la publicación de este artículo.
FINANCIAMIENTO
Autofinanciado.
AGRADECIMIENTO
A todos los agentes sociales involucrados en el proceso investigativo.
REFERENCIAS
1. Borland JM, Frantz KJ, Aiani LM, Grantham KN, Song Z, Albers HE. A novel operant task to assess social reward and motivation in rodents. J Neurosci Methods. 2017;287:80-88. http://dx.doi.org/10.1016/j.jneumeth.2017.06.003
2. Song Z, Borland JM, Larkin TE, O'Malley M, Albers HE. Activation of oxytocin receptors, but not arginine-vasopressin V1a receptors, in the ventral tegmental area of male Syrian hamsters is essential for the reward-like properties of social interactions. Psychoneuroendocrinology. 2016;74:164-172. http://dx.doi.org/10.1016/j.psyneuen.2016.09.001
3. Borland JM, Aiani LM, Norvelle A, et al. Sex-dependent regulation of social reward by oxytocin receptors in the ventral tegmental area. Neuropsychopharmacology. 2019;44(4):785-792. http://dx.doi.org/10.1038/s41386-018-0262-y
4. Clinard CT, Bader LR, Sullivan MA, Cooper MA. Activation of 5-HT2a receptors in the basolateral amygdala promotes defeat-induced anxiety and the acquisition of conditioned defeat in Syrian hamsters. Neuropharmacology. 2015;90:102-112. http://dx.doi.org/10.1016/j.neuropharm.2014.11.016
5. González-Martínez LF, D'Aigle J, Lee SM, Lee HJ, Delville Y. Social stress in early puberty has long-term impacts on impulsive action. Behav Neurosci. 2017;131(3):249-261. http://dx.doi.org/10.1037/bne0000196
6. Borland JM, Grantham KN, Aiani LM, Frantz KJ, Albers HE. Role of oxytocin in the ventral tegmental area in social reinforcement. Psychoneuroendocrinology. 2018;95:128-137. http://dx.doi.org/10.1016/j.psyneuen.2018.05.028
7. Miler K, Scharf I. Operant conditioning in antlion larvae and its impairment following exposure to elevated temperatures. Anim Cogn. 2022;25(3):509-518. http://dx.doi.org/10.1007/s10071-021-01570-9
8. Guttlein L, Molina JC, Abate P. Operant conditioning with a stimulus discrimination: An alternative method for evaluating alcohol reinforcement in preweaning rats. J Neurosci Methods. 2021;363:109345. http://dx.doi.org/10.1016/j.jneumeth.2021.109345
9. Eftekhar A, Norton JJS, McDonough CM, Wolpaw JR. Retraining Reflexes: Clinical Translation of Spinal Reflex Operant Conditioning. Neurotherapeutics. 2018;15(3):669-683. http://dx.doi.org/10.1007/s13311-018-0643-2
10. Adelina N, Chiu CHM, Lam K, Takano K, Barry TJ. Social operant conditioning of autobiographical memory sharing. Behav Res Ther. 2023;168:104385. http://dx.doi.org/10.1016/j.brat.2023.104385
11. Eder AB, Krishna A, Van Dessel P. Operant evaluative conditioning. J Exp Psychol Anim Learn Cogn. 2019;45(1):102-110. http://dx.doi.org/10.1037/xan0000189
12. Bąbel P. Operant conditioning as a new mechanism of placebo effects. Eur J Pain. 2020;24(5):902-908. http://dx.doi.org/10.1002/ejp.1544
13. Chikamoto N, Fujimoto K, Nakai J, Namiki K, Hatakeyama D, Ito E. Genes Upregulated by Operant Conditioning of Escape Behavior in the Pond Snail Lymnaea stagnalis. Zoolog Sci. 2023;40(5):375-381. http://dx.doi.org/10.2108/zs230032
14. Lintas A, Sánchez-Campusano R, Villa AEP, Gruart A, Delgado-García JM. Operant conditioning deficits and modified local field potential activities in parvalbumin-deficient mice. Sci Rep. 2021;11(1):2970. http://dx.doi.org/10.1038/s41598-021-82519-3
15. Ito H, Fujiki S, Mori Y, Kansaku K. Self-reorganization of neuronal activation patterns in the cortex under brain-machine interface and neural operant conditioning. Neurosci Res. 2020;156:279-292. http://dx.doi.org/10.1016/j.neures.2020.03.008
©2024 por los autores. Este artículo es de acceso abierto y distribuido según los términos y condiciones de la licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional (CC BY-NC-SA 4.0) (https://creativecommons.org/licenses/by-nc-sa/4.0/).